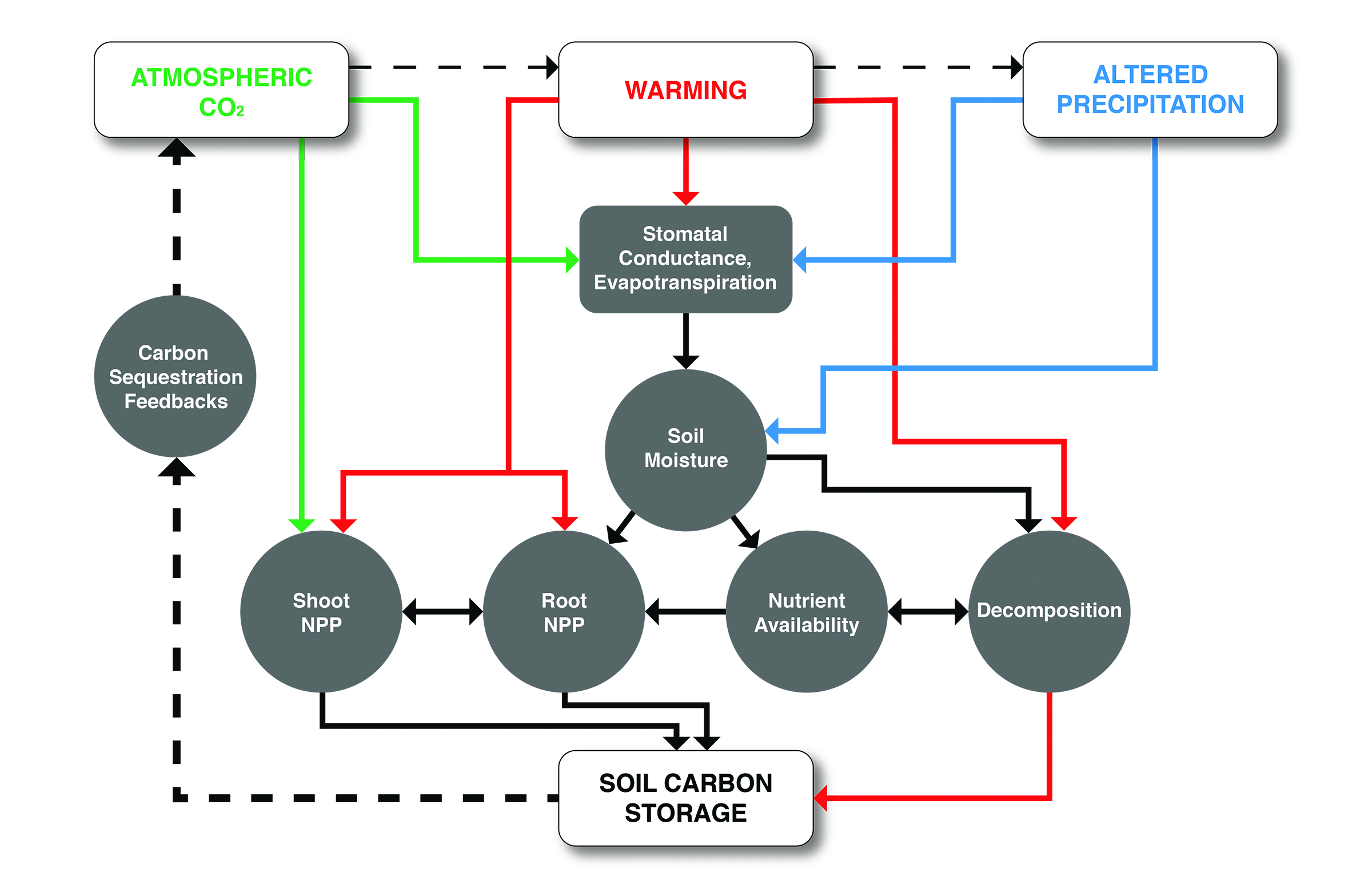
Key Finding 1
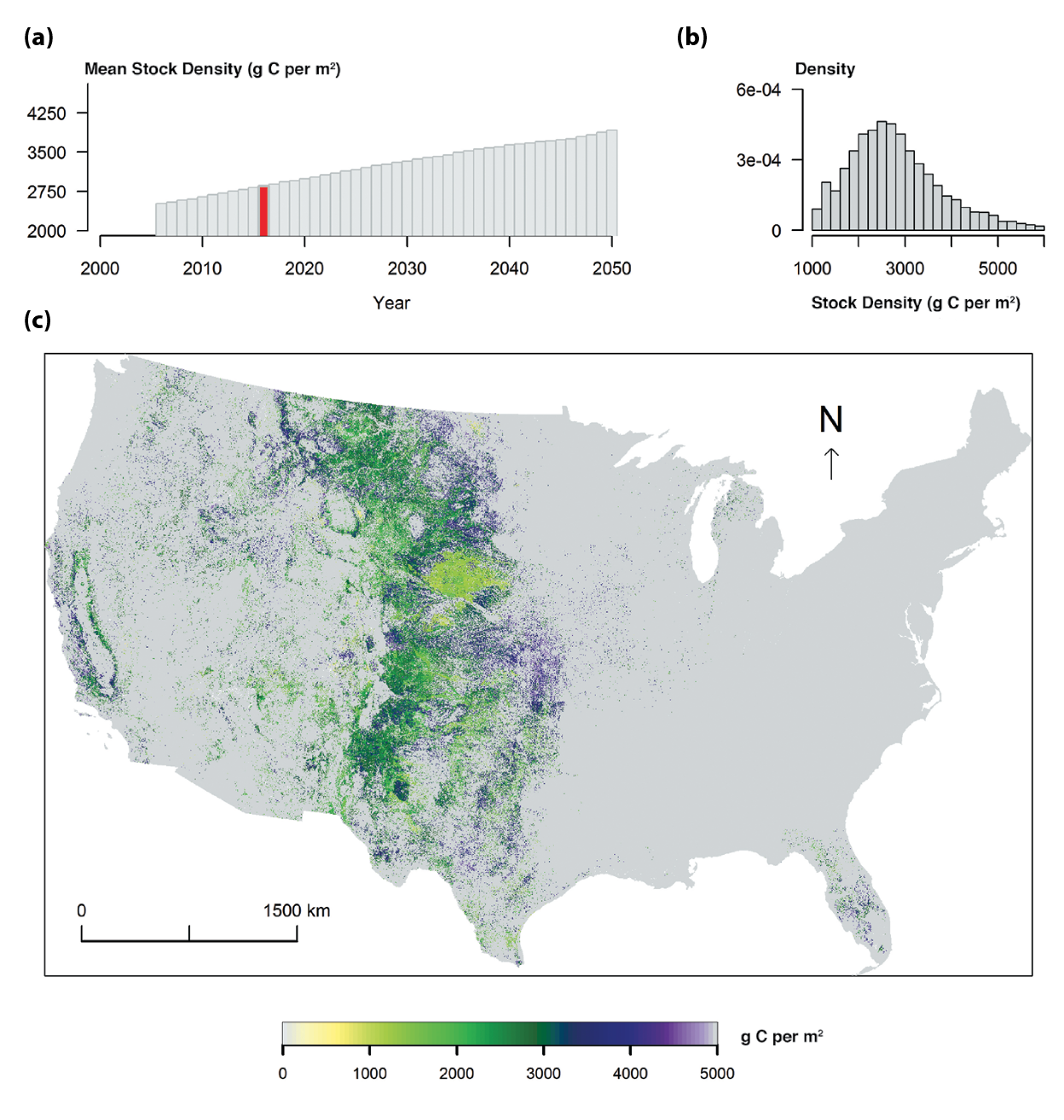
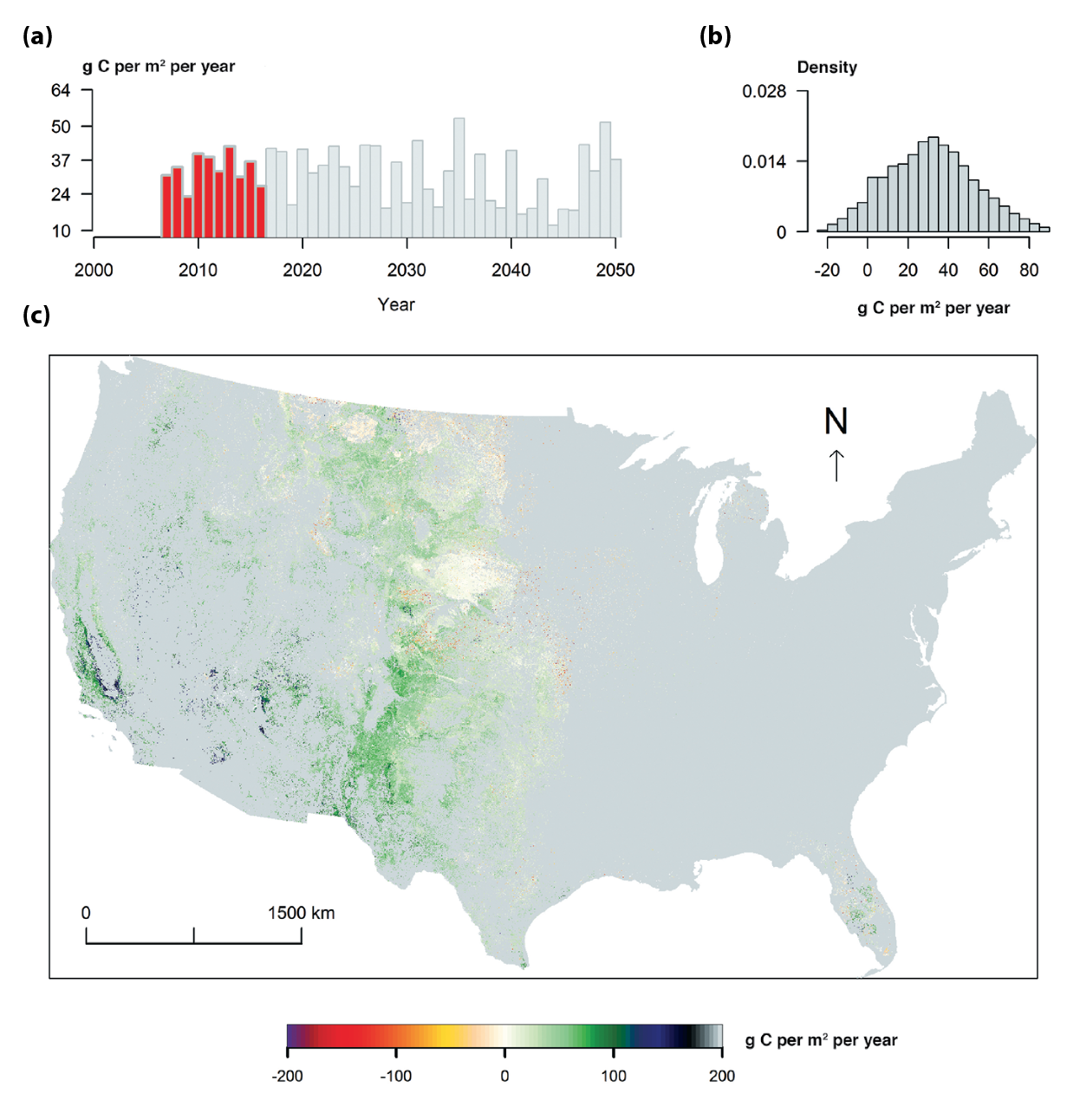
Total grassland carbon stocks in the conterminous United States, estimated to be about 7.4 petagrams of carbon (Pg C) in 2005, are projected to increase to about 8.2 Pg C by 2050. Although U.S. grasslands are expected to remain carbon sinks over this period, the uptake rate is projected to decline by about half. In the U.S. Great Plains, land-use and land-cover changes are expected to cause much of the change in carbon cycling as grasslands are converted to agricultural lands or to woody biomes (medium confidence).
Description of evidence base
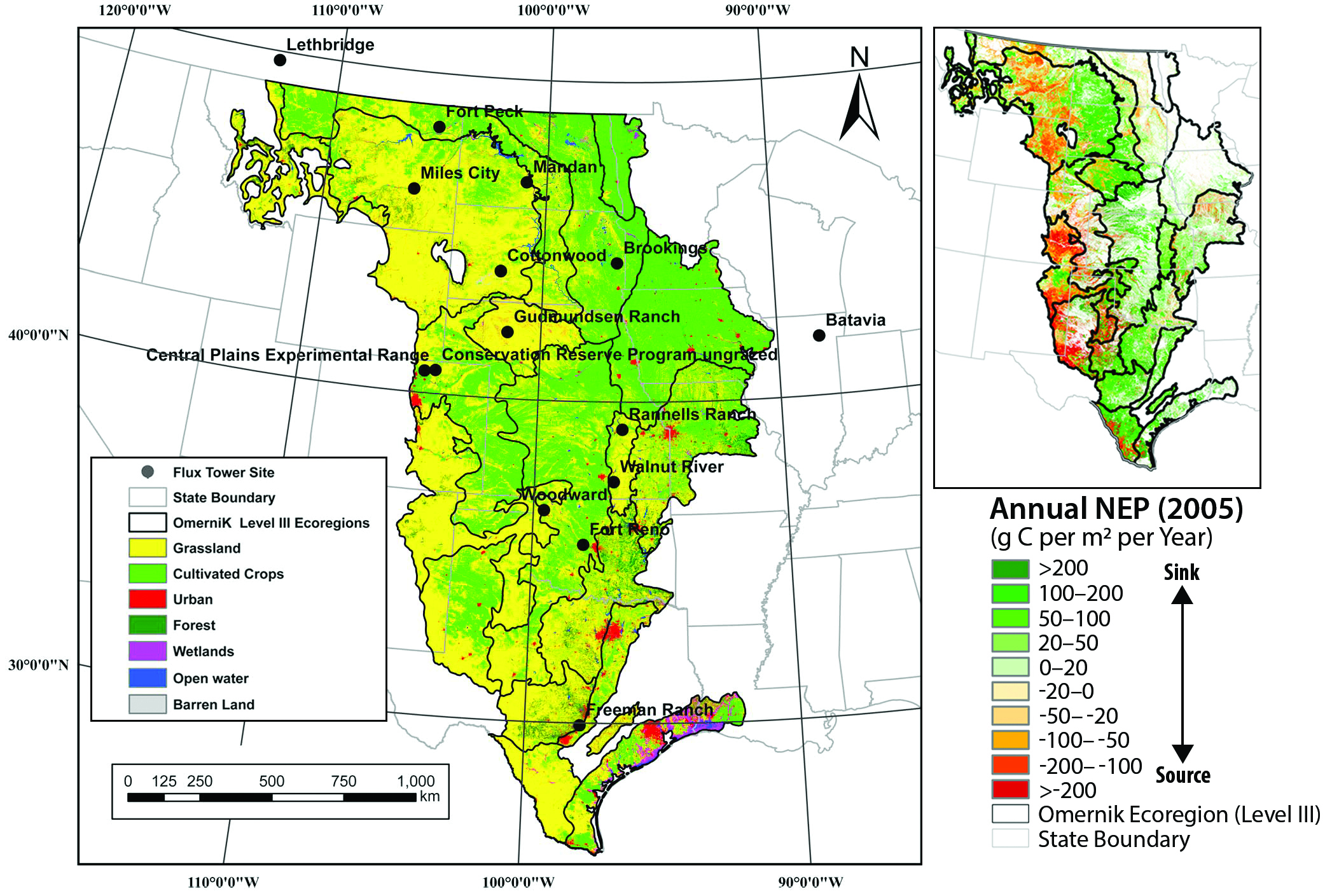
Total carbon stocks are from Table 10.2 based on LandCarbon project estimates (landcarbon.org/categories). Various efforts confirm that the U.S. and North American grasslands in recent years have been a weak carbon sink (i.e., mostly within the range of 10 to 40 g per m2 per year; Hayes et al., 2012; Liu et al., 2012b; Raczka et al., 2013; Wylie et al., 2016; Xiao et al., 2014; Zhang et al., 2011). Recent results generated from the assessment of carbon sequestration potentials in the United States conducted by the U.S. Geological Survey (Zhu and Reed 2012, 2014; Zhu et al., 2011) provided more integrated grassland carbon assessment. Land-use change scenarios and spatial dynamics were developed empirically by ecoregions across the United States under the Intergovernmental Panel on Climate Change (IPCC) scenarios A1B, A2, and B1 (Sleeter et al., 2012; Sohl et al., 2007), which are considered to be similar to representative concentration pathway (RCP) scenarios (Knutti and Sedláček 2013). Carbon dynamics in grassland ecosystems were simulated with the General Ensemble Biogeochemical Modeling System (GEMS) using three climate projections: the Second Generation Coupled Global Climate Model (CGCM2), Australia’s national Commonwealth Science and Industry Research Organization (CSIRO), and Model for Interdisciplinary Research on Climate (MIROC) for each of the three IPCC scenarios (Liu et al., 2012b, 2014). The data included in this report include simulations from two process-based models: CENTURY (Parton et al., 1987) and the Erosion-Deposition–Carbon-Model (EDCM; Liu et al., 2003), and both were encapsulated in GEMS. The findings are supported by a recent synthesis of eddy covariance data with remote sensing, which shows that grasslands take up somewhat more carbon than crops in the Great Plains, although both were weak carbon sinks from 2000 to 2008 (Wylie et al., 2016).
Major uncertainties
There are significant differences in evaluation of grassland carbon stocks and fluxes (Hayes et al., 2012; Raczka et al., 2013; Zhu and Reed 2014). The primary source of model difference comprises modeling method (i.e., inventory, flux towers, inversion, and process-based modeling) and land-cover characterization and spatial resolution. For example, the LandCarbon study (Zhu and Reed 2012, 2014; Zhu et al., 2011) combined grass and shrub into grassland and considered fire disturbance, while Zhang et al. (2011) used data from 15 flux towers at natural grassland and pastures or hay sites but without considering fires.
Assessment of confidence based on evidence and agreement, including short description of nature of evidence and level of agreement
The magnitudes of the estimates of carbon stocks and fluxes vary depending on the method used, indicating a medium to low level of confidence in the results.
Summary sentence or paragraph that integrates the above information
Grasslands appear very likely to be weak carbon sinks and will remain so for at least the coming three decades, but reconciling different methods will reduce uncertainties in the quantities.
Key Finding 2
Increasing temperatures and rising atmospheric carbon dioxide (CO2) concentrations interact to increase productivity in northern North American grasslands, but this productivity response will be mediated by variable precipitation, soil moisture, and nutrient availability (high confidence, very likely).
Description of evidence base
Experimental manipulations in the field provide evidence of climate change effects on grassland productivity by up to 33%, but this is contingent on nutrient and moisture availability (e.g., Morgan et al., 2011; Mueller et al., 2016; Reich and Hobbie 2013). Spatially distributed observations of vegetation phenology (i.e., greenness) and carbon fluxes combined with empirical modeling provide evidence of regional differences in grassland responses to future climate change (Hufkens et al., 2016). Simulation models are in general agreement with empirical evidence that carbon stocks will increase in grasslands in the coming three to four decades (Zhu et al., 2011). In grasslands, carbon uptake is responsive to precipitation amount and event size and timing, with both positive and negative effects documented, but droughts are associated with carbon losses across all grasslands (Cherwin and Knapp 2012; Goldstein and Suding 2014; Heisler-White et al., 2008, 2009; Knapp et al., 2008b; Kulmatiski and Beard 2013; Thomey et al., 2011).
Major uncertainties
The largest source of uncertainty is related to future precipitation regimes in the grassland biomes of North America, with both increases and decreases in precipitation predicted (IPCC 2013). The degree to which altered precipitation regimes will affect carbon cycling in future grasslands is uncertain (Knapp et al., 2008b). The relative response of grassland productivity to moisture availability is contingent upon prior conditions, which vary temporally and spatially (Heisler-White et al., 2009). Empirical models represent grassland phenology and productivity well, but they lack explicit physiological processes, leading to uncertainties in mechanisms underlying ecosystem responses to climate change (Hufkens et al., 2016).
Assessment of confidence based on evidence and agreement, including short description of nature of evidence and level of agreement
Confidence is high that grassland production will increase with precipitation as atmospheric CO2 and temperature increase in the coming three to four decades, based on empirical evidence from field experiments.
Estimated likelihood of impact or consequence, including short description of basis of estimate
If grassland productivity decreases in response to climate change, such as reduced precipitation, forage production for livestock is very likely to be at risk. This has been demonstrated by numerous experiments and models as explained above in the description of evidence base.
Summary sentence or paragraph that integrates the above information
Grassland productivity is highly likely to respond positively to increased precipitation and temperature, especially in the Northern Great Plains. Neutral or negative responses of productivity to warming in the Southern Great Plains, the southwestern United States, and Mexico may be offset by positive responses to elevated CO2.
Key Finding 3
Soil carbon in grasslands is likely to be moderately responsive to changes in climate over the next several decades. Field experiments in grasslands suggest that altered precipitation can increase soil carbon, while warming and elevated CO2 may have only minimal effects despite altered productivity (medium confidence, likely).
Description of evidence base
Meta-analysis of numerous field experiments showed that soil carbon stocks increase when precipitation is increased or decreased in grasslands (Zhou et al., 2016). Meta-analysis also showed that elevated CO2 increased soil carbon decomposition rate, limiting carbon storage potential (van Groenigen et al., 2014). Field experiments indicate that soil carbon stocks decrease with warming, especially in regions where stocks are high to begin with (Crowther et al., 2016), although warming-induced soil carbon losses from grasslands may be insignificant (Lu et al., 2013). These results are confirmed in some simulation experiments (e.g., Parton et al., 2007; Shi et al., 2015).
Major uncertainties
Major uncertainties in soil carbon storage come from insufficient understanding of physical and biological mechanisms that determine the stability of soil carbon. Physical mechanisms underlying carbon stability in soil, such as protection within aggregates and their sensitivity to climate change, are still poorly described (Heimann and Reichstein 2008). In particular, regulation of soil organic matter decomposition by microbe-plant interactions is poorly understood and not well represented in models (Wieder et al., 2015). Improving mechanistic understanding of soil carbon dynamics, and incorporating key mechanisms into models, will reduce uncertainties in future carbon cycle predictions (Todd-Brown et al., 2013).
Assessment of confidence based on evidence and agreement, including short description of nature of evidence and level of agreement
Mechanistic understanding of soil carbon stability in the face of climate change is still limited, leading to only medium confidence levels regarding the response of soil carbon to climate changes.
Estimated likelihood of impact or consequence, including short description of basis of estimate
Soils in grasslands are not likely to respond strongly to climate change; small carbon losses or gains could occur in the future with warming or elevated CO2. Larger carbon gains are likely to occur with increased precipitation.
Summary sentence or paragraph that integrates the above information
Mechanisms regulating soil carbon storage in response to climate change can be incorporated into models to improve confidence in model predictions of future carbon cycling.
Key Finding 4
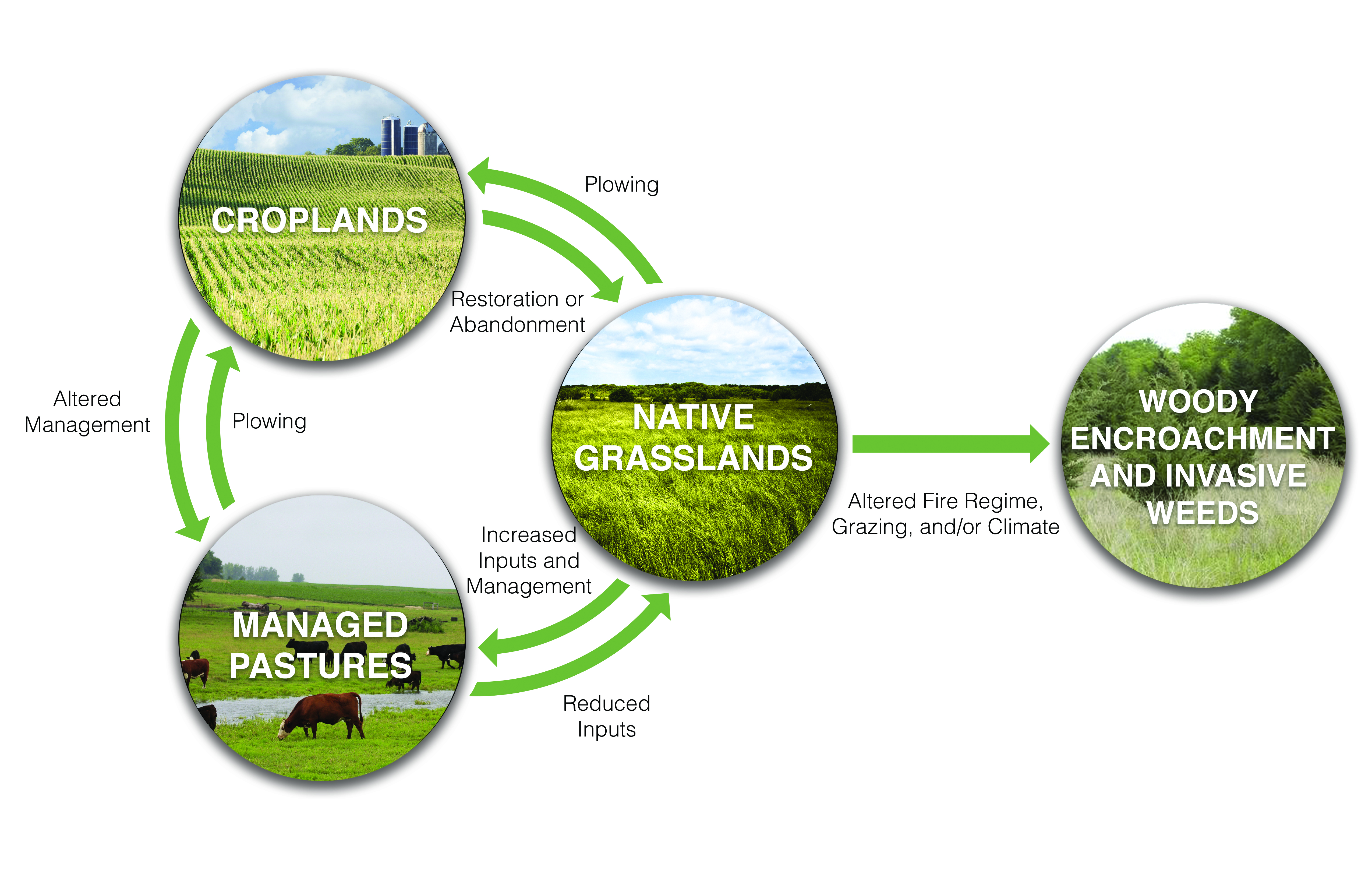
Carbon stocks and net carbon uptake in grasslands can be maintained with appropriate land management including moderate levels of grazing. Fire suppression can lead to encroachment of woody vegetation and increasing carbon storage in mesic regions, at the expense of grassland vegetation (high confidence, likely).
Description of evidence base
Studies of carbon fluxes using eddy covariance indicate that moderate grazing allows grasslands to continue to be net carbon sinks, but heavy grazing diminishes their capacity to take up carbon (Frank 2004; Morgan et al., 2016; Polley et al., 2008; Risch and Frank 2006). Soil inventory studies indicate that moderate to light grazing does not negatively affect carbon stocks (Conant et al., 2001, 2017), and improving grazing management can augment carbon stocks (Chambers et al., 2016). Carbon cycle responses to woody encroachment are determined from inventories of carbon stocks in vegetation and soils in plots that have been experiencing woody encroachment for different periods of time (Barger et al., 2011; Knapp et al., 2008a).
Major uncertainties
Uncertainties in grazing management impacts on carbon cycling in grasslands stem mainly from the regional variations in soil carbon responses to management, from challenges in designing scientific studies that adequately represent real-world management practices, and from limitations faced when extrapolating plot-level studies to broader areas (Conant et al., 2017). Interactive effects of grazing, climate, soil type and plant community composition on carbon storage are not well constrained (McSherry and Ritchie 2013). The magnitude of carbon accumulation below ground in response to woody encroachment is poorly constrained, but change in carbon pools above ground is well known (Barger et al., 2011; Knapp et al., 2008a). Fire regimes are changing with increasing temperatures and altered vegetation; uncertainties in future fire risk add uncertainty to projections of carbon budgets.
Assessment of confidence based on evidence and agreement, including short description of nature of evidence and level of agreement
There is high confidence with general agreement across several studies that moderate to light grazing will not have a negative impact on carbon cycling.
Estimated likelihood of impact or consequence, including short description of basis of estimate
Woody encroachment likely will lead to increased carbon storage in mesic grasslands.
Summary sentence or paragraph that integrates the above information
Carbon likely will continue to accumulate for the next several decades in grasslands if they are appropriately managed.
Supporting Evidence for Tables
Table 10.1 is based on Hayes et al. (2012). The areas for grasslands by countries and the continent are from the models and inventory analyses used in their study (see Table S10 in Hayes et al., 2012). The area for “Others” is smaller for the models than the inventory analysis mainly because the latter includes urban areas. Inventory estimates are the sum of livestock methane (CH4) emissions + livestock carbon dioxide (CO2) emissions + grassland net ecosystem exchange (NEE) for Canada and the United States. Taiga was excluded from Canada grassland NEE and livestock emissions. For Mexico, the number for “Others” was used because extracting grassland NEE was not possible. Atmospheric inversion models (AIMs) and land-surface models (LSMs) are from Table 2 in Hayes et al. (2012) and do not include CH4 emissions or human settlement emissions. Thus, the AIM values of NEE for “Others” should be representative of grassland and pastureland NEE. Area estimate for grasslands: www.statista.com/statistics/201761/projection-for-total-us-grassland-area-from-2010.
Table 10.2 Carbon fluxes and stocks for grasslands and shrublands in the conterminous United States summarized from the LandCarbon project (landcarbon.org/categories). Values are averages of the A1B, A2, and B1 climate scenarios and estimated using the FOREcasting SCEnarios of land-use change (FORE-SCE) model and the Erosion-Deposition-Carbon-Model (EDCM), CENTURY, and PBN carbon models (Liu et al., 2012b, 2014; Zhu et al., 2011). Climate projections based on emissions scenarios used by the LandCarbon Project are considered to be similar to representative concentration pathway (RCP) scenarios (Knutti and Sedláček 2013). Negative fluxes indicate carbon losses from the ecosystem; positive fluxes indicate carbon gains by the ecosystem. The total flux is considered to be the net ecosystem carbon balance (NECB). Land-cover classification could be a source of differences. Flux towers mostly measure actual grassland and rangeland, whereas the General Ensemble Biogeochemical Modeling System (GEMS) includes both grassland and shrubland. The conterminous United States has about 1 million km2 of grassland and 1.3 million km2 of shrubland (from Liu et al. land-cover data). The area difference is notable. Land conversion to and from agriculture and permanent grassland loss to urban land all contribute to the total carbon number.