<b>Windham-Myers</b>, L., W.-J. <b>Cai</b>, S. R. Alin, A. Andersson, J. Crosswell, K. H. Dunton, J. M. Hernandez-Ayon, M. Herrmann, A. L. Hinson, C. S. Hopkinson, J. Howard, X. Hu, S. H. Knox, K. Kroeger, D. Lagomasino, P. Megonigal, R. G. Najjar, M.-L. Paulsen, D. Peteet, E. Pidgeon, K. V. R. Schäfer, M. Tzortziou, Z. A. Wang, and E. B. Watson, 2018: Chapter 15: Tidal wetlands and estuaries. In Second State of the Carbon Cycle Report (SOCCR2): A Sustained Assessment Report [Cavallaro, N., G. Shrestha, R. Birdsey, M. A. Mayes, R. G. Najjar, S. C. Reed, P. Romero-Lankao, and Z. Zhu (eds.)]. U.S. Global Change Research Program, Washington, DC, USA, pp. 596-648, https://doi.org/10.7930/SOCCR2.2018.Ch15.
Tidal Wetlands and Estuaries
Tidal wetlands and estuaries of North America vary in relative area depending on coastal topography, historic rates of sea level rise, and inputs of suspended solids from land. In drowned river valleys (e.g., Chesapeake Bay) and fjords (e.g., Puget Sound) that are topographically steep, estuarine habitat is the dominant subsystem (Dalrymple et al., 1992). In contrast, the ratio of tidal wetland area to estuarine area is relatively high (Day et al., 2013), though still less than one (Najjar et al., 2018) along coastal plains.
The land-sea interface that defines the presence of tidal wetlands and estuaries (i.e., river-sea mixing zones) is itself extremely dynamic over broad spatial and temporal scales. The current configuration of tidal wetlands and estuaries is the result of processes that have been occurring since the last glacial maximum, roughly 18,000 years ago. Over the past 6,000 years, when rates of sea level rise slowed to less than 1 mm per year, tidal wetlands increased in size relative to open-water estuaries, as bay bottoms filled with sediments from uplands and tidal wetlands prograded into shallow open-water regions and transgressed across uplands (see Figure 15.1; Redfield 1967). Concomitant with increasing sea levels, tidal wetlands maintained their relative elevation as wetland plants trapped suspended sediments from tidal floodwaters, as well as accumulated organic matter in soils. Factors that affect tidal wetland area and relative elevation, through lateral and vertical erosion and accretion, include 1) rate of sea level rise, 2) land subsidence or isostasy (glacial rebound), 3) delivery and deposition of suspended sediment, 4) balance between wetland gross primary production (GPP) and respiration of all autotrophs and heterotrophs (RAH), 5) sediment compaction, and 6) slope of land at the land-water interface (Cahoon 2006).
Figure 15.1: Conceptual Model of Coastal Tidal Wetlands and Estuaries and Their Linkages with Adjacent Terrestrial and Oceanic Systems
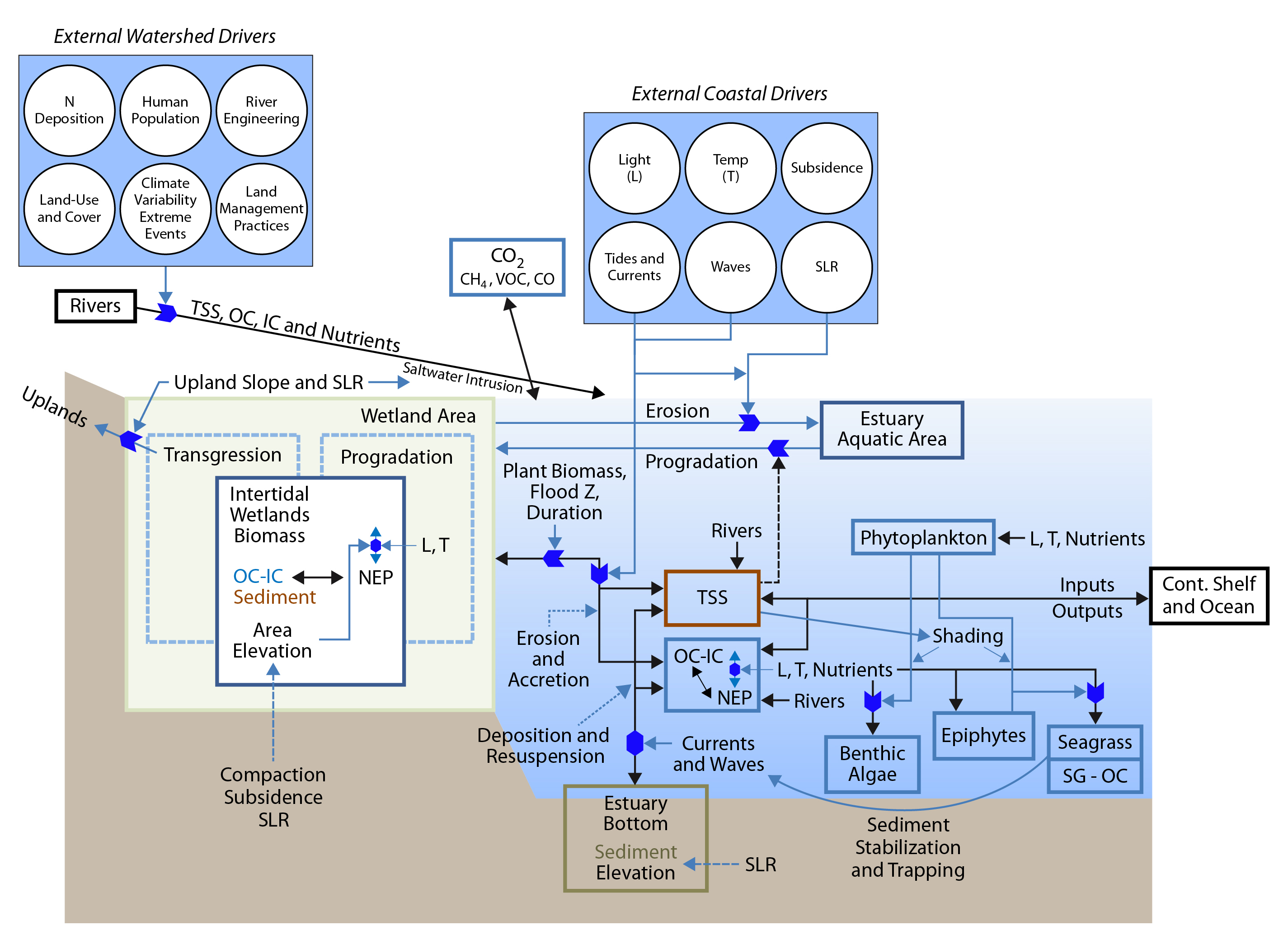
Tidal wetlands are among the most productive ecosystems on Earth, continuously accumulating organic carbon that results from environmental conditions that inhibit organic matter decomposition. As a result, intact tidal wetlands are capable of storing vast amounts of autochthonous organic carbon (i.e., fixed through photosynthesis on site) as well as intercepting and storing allochthonous organic carbon (i.e., produced off site, terrigenous; Canuel et al., 2012). Documented carbon-related ecosystem benefits, referred to as “services,” include significant uptake and storage of carbon in wetland soils, as well as export to the ocean of organic matter, which increases the productivity of coastal fisheries (Day et al., 2013). Globally, tidal wetlands are strongly variable in age and structure. Some of today’s tidal wetlands have persisted for more than 6,500 years, accumulating to a depth of up to 13 m of tidal peat (Drexler et al., 2009; McKee et al., 2007; Peteet et al., 2006), but some wetlands are young and shallow because of recent human influences that enhanced sediment delivery to nearshore waters. Examples include the colonial-era East Coast (Kirwan et al., 2011) and gold rush in California (Palaima 2012). Because human development is preferentially concentrated on coastlines, tidal wetlands have been subject to active loss through development pressures. While tidal wetland losses have slowed in the United States, global tidal wetland losses are currently estimated at 0.5% to 3% annually (Pendleton et al., 2012), with estimates depending on the ecosystem, time frame, and methods used in evaluation (Hamilton and Casey 2016; Spalding et al., 2010). Loss of carbon stocks through wetland drainage and erosion remains poorly modeled due to limited mapping and quantification of initial carbon stock conditions (Chmura 2013). Further, more subtle rates of wetland loss, through drowning or erosion, may be underestimated by remote-sensing techniques insensitive to small-scale changes observed through aerial photography (e.g., Schepers et al., 2017; Watson et al., 2017).
Estuarine waters are a small but productive fraction of coastal waters (Cloern et al., 2014; Wollast 1991). The role of coastal zones as sinks or sources of atmospheric CO2 is still poorly understood (Borges 2005; Borges et al., 2005; Smith and Hollibaugh 1997), resulting in a lack of consensus toward their role in global carbon budgets (Cai 2011; Wollast 1991; Borges and Abril 2011; Chen et al., 2013). With poorly characterized boundary conditions, estuarine waters have strong upland and ocean-based drivers, leading to strong seasonality in carbon transport and transformation. Geological records suggest that estuarine carbon storage was enhanced in the past 6,000 years and during recent centuries by watershed activities (Colman et al., 2002), but responses were varied. Human activities initially increased the delivery of organic materials to estuaries (e.g., forest clearing) and thus drove them to support higher net respiration (and likely greater sources of atmospheric CO2); however, more recent human activities (e.g., dam construction and fertilizer use) have greatly reduced sediment and organic matter delivery but increased nutrient fluxes to many estuaries (Bianchi and Allison 2009; Galloway et al., 2008), driving estuarine waters to be less heterotrophic and, possibly, causing more net carbon burial and export to the ocean (Regnier et al., 2013). While North American estuarine conditions vary along coasts according to upstream land use, the most significant human-induced change to estuarine carbon dynamics over the past century is certainly increased nutrient loading (Schlesinger 2009), which has led to eutrophication and hypoxia in estuaries and continental shelves. Eutrophication promotes carbon uptake and pH increase in surface estuarine waters (Borges and Gypens 2010), but it also may enhance acidification when organic matter fixed by photosynthesis is respired. In stratified estuarine waters, respiration-induced CO2 and poor buffering capacity could greatly reduce pH and carbonate saturation states to levels much lower than those resulting from the increase of anthropogenic CO2 in the atmosphere and its subsequent uptake in surface waters (Cai 2011, Cai et al., 2017; Feely et al., 2010). The particularly large pH changes and the difficulty in predicting acidification in estuaries have motivated many scientists to study estuarine acidification in addition to ocean acidification (Duarte et al., 2013).
Estuaries generally have more interannual variability in carbon dynamics than do tidal wetlands, a phenomenon reflecting the balance of exchanges with terrestrial watersheds, tidal wetlands, and the continental shelf (Bauer et al., 2013). Processing of material inputs from land and tidal wetlands determines the autotrophic-heterotrophic balance of the estuary; this processing reflects the biological, chemical, and physical structure of the receiving estuary, as well as the nature of the inputs themselves. The autotrophic-heterotrophic balance of an estuary is especially sensitive to the water residence time (largely a function of freshwater runoff, tidal mixing, and estuarine geometry), the ratio of inputs of organic carbon (primarily from land and tidal wetlands) to inorganic nutrients (primarily from land), the degradability of the organic carbon input (Hopkinson and Vallino 1995; Kemp et al., 1997; Herrmann et al., 2015). The relative abundance of pelagic (i.e., phytoplankton-dominated) versus benthic (i.e., seagrass- or benthic algal–dominated) communities is also a major factor affecting estuarine carbon dynamics. The availability of light is perhaps the major constraint on the distribution of benthic autotrophic communities. Light availability to the benthos depends on estuarine depth and water clarity, which in turn are related to concentrations of suspended solids and phytoplankton in the estuarine water column. In nitrogen-enriched estuarine waters, high-phytoplankton biomass and epiphytic algae decrease light availability to benthic autotrophic communities, sometimes resulting in a complete loss of seagrass habitats (Howarth et al., 2000). In shallow systems, benthic macroalgae often dominate system dynamics. Seagrass, because of its ability to control wave and current strength, can play a major role in limiting sediment resuspension, thereby maintaining high water clarity (van der Heide et al., 2011). Estuaries typically are heterotrophic and release CO2 to the atmosphere, largely as a result of their processing of organic carbon inputs from watersheds (Raymond and Bauer 2001) and adjacent tidal wetlands (Bauer et al., 2013; Cai and Wang 1998; Wang and Cai 2004). For example, U.S. Atlantic coastal estuaries as a whole are net heterotrophic (Herrmann et al., 2015); all but three of 42 sites in the U.S. National Estuarine Research Reserve System were net heterotrophic over a year (Caffrey 2004), and a global survey concluded that 66 out of 79 estuaries were net heterotrophic (Borges and Abril 2011). At the same time, estuaries can serve as significant long-term organic carbon sinks through sedimentation of terrestrial inputs and seagrass organic matter burial (Duarte et al., 2005; Hopkinson et al., 2012; McLeod et al., 2011; Nellemann et al., 2009).
See Full Chapter & References