<b>Pendall</b>, E., D. Bachelet, R. T. Conant, B. El Masri, L. B. Flanagan, A. K. Knapp, J. Liu, S. Liu, and S. M. Schaeffer, 2018: Chapter 10: Grasslands. In Second State of the Carbon Cycle Report (SOCCR2): A Sustained Assessment Report [Cavallaro, N., G. Shrestha, R. Birdsey, M. A. Mayes, R. G. Najjar, S. C. Reed, P. Romero-Lankao, and Z. Zhu (eds.)]. U.S. Global Change Research Program, Washington, DC, USA, pp. 399-427, https://doi.org/10.7930/SOCCR2.2018.Ch10.
Grasslands
10.3.1 Future Projections of Carbon Stocks and Fluxes in Conterminous U.S. Grasslands
In estimating carbon stock and fluxes, several different models were used (see Key Finding 1) to assess their projections, The LandCarbon project simulated future carbon stocks (see Figure 10.2) and fluxes (see Figure 10.3) using projections from MIROC A1B, A2, and B1 climate scenarios; FORE-SCE model; and EDCM (Liu et al., 2012b, 2014). Thus, these simulations combine the effects of land-use change and climate on carbon sequestration by grasslands in the conterminous United States (see Table 10.2). While these model predictions are useful as general guidelines, additional empirical and simulation experiments are needed to disaggregate the effects of land-cover change from those of climate change and to examine regional differences in carbon cycling.
10.3.2 Impacts of Land-Use and Land-Cover Change on Future Carbon Cycling
Zhu et al. (2011) demonstrate that land-use and land-cover conversions were major drivers of the predicted changes in carbon storage in Great Plains grasslands. Future land-use change in the region (data provided by the Intergovernmental Panel on Climate Change’s Special Report on Emission Scenarios; IPCC 2000) is driven by the demand for agricultural commodities, including biofuels, resulting in a 1.4% to 9.2% expansion of agricultural land by 2050, mostly at the expense of grasslands (–2.2% to –9.3%). Areas where woody vegetation expands into grassland because of fire suppression are re-classified as forest. This change tends to result in higher carbon stocks and uptake rates but also can be subject to catastrophic carbon losses in hot and dry fire years following wet years’ boosting of fuel loads (Bachelet et al., 2017).
In the Great Plains, carbon stocks for the years 2001 to 2005 are assessed as 7,500 Tg C with 45.8% in agricultural lands, 34.9% in grasslands and shrublands, 15.5% in the few existing forested areas, and almost 3% in wetlands. By 2050, models estimate those percentages will change to reflect a small increase in agricultural land carbon stocks (47%), a large decrease in grassland carbon stocks (29%), an increase in forestland carbon stocks (20.4%) due to woody encroachment and forest growth, and no change in carbon stocks of wetlands or other lands. Conversion of grasslands to agriculture may lead to a cumulative reduction in stored carbon of 26 to 157 Tg from 2001 to 2050, an amount which could contribute up to 4% loss of mean total carbon sequestration potential (Zhu et al., 2011). Shrub encroachment and afforestation cannot mitigate carbon losses to agricultural expansion. Fires are also a source of carbon loss. Areas burned and carbon emissions from fires vary both spatially and temporally due to climatic, biological, and physical factors. However, fires in grasslands were not projected to change significantly under future climate conditions when models did not include the role of annual invasives or fire suppression. Average fire emissions from grasslands range from 0.18 to 24.72 Tg CO2 equivalent1 (CO2e) per year (Zhu et al., 2011).
10.3.3 Climate Change Impacts on Grassland Productivity
Numerous environmental factors interact to affect grassland production, including warming, rising CO2, hydrology, and nutrient availability. Grassland productivity is very sensitive to variations in climate, especially precipitation and including both the mean and extremes such as droughts and floods (Huxman et al., 2004; Knapp et al., 2001, 2008b, 2015). Their sensitivity indicates a strong potential for climate change to alter carbon cycling in grasslands (see Key Finding 2; Figure 10.5). Productivity is predicted to decline in the southwestern United States and northern Mexico as a result of reduced precipitation and to increase in the northern Great Plains as a result of temperature and precipitation increases that allow an increase in growing season length (Hufkens et al., 2016; Polley et al., 2013; Reeves et al., 2014). However, significant projected increases in productivity did not arise until after 2030 because of scenarios projecting CO2 fertilization and rising temperatures (Reeves et al., 2014).
Figure 10.5: Interacting Effects of Rising Atmospheric Carbon Dioxide (CO2), Warming, and Altered Precipitation on Grasslands
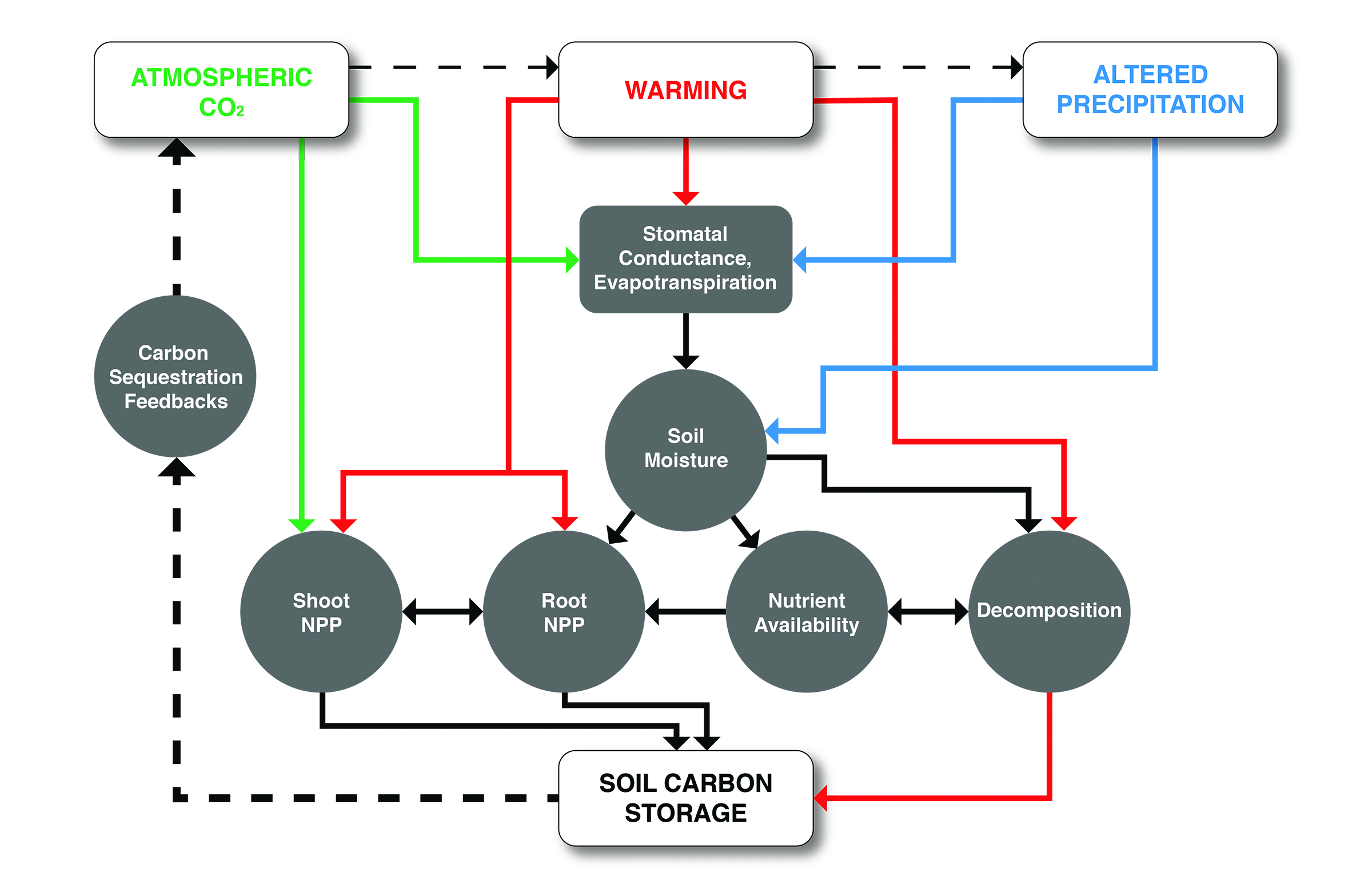
North American grassland growth in this century was simulated based on hydrology and repeat-photography observations of vegetation greenness (Hufkens et al., 2016). Despite a projected increase in climate aridity by 2100, increases in fractional plant cover were predicted over almost 90% of the study area, with greater increases in cover and net carbon sequestration in the more northerly areas. The primary mechanism contributing to the projected increase in grassland growth was a shift to earlier leaf emergence in the spring and delayed leaf senescence in the autumn, both of which compensated for drought-induced reduction in plant productivity during the summer (Hufkens et al., 2016).
Predictions from the vegetation-hydrology model are supported by a climate manipulation experiment in Wyoming mixed-grass prairie, where the growing season started earlier in spring because of the warming treatment and ended later in autumn because of increased soil moisture made available by the elevated CO2 treatment (Reyes-Fox et al., 2014). The lengthening of the growing season was dependent on a mix of C3 and C4 species adapted to different climate conditions. In the same experiment, greenness was enhanced (i.e., indicating increased aboveground biomass and cover) with warming and elevated CO2, but the effects of seasonal and interannual rainfall variability were much stronger (Zelikova et al., 2015). High-precipitation years had two to three times greater vegetation greenness than dry years. Warming in combination with elevated CO2 increased total plant biomass by an average of 25%, especially below ground (Mueller et al., 2016). Warming and elevated CO2 also interacted to affect soil moisture and nitrogen availability (Mueller et al., 2016). While elevated CO2 conditions increased soil moisture (Morgan et al., 2011), warming decreased soil moisture, and soil nitrate tended to follow trends opposite to those for elevated CO2 (Mueller et al., 2016). A warming experiment in desert grasslands suggested warming could reduce C3 and C4 grass carbon fixation rates and aboveground biomass, with no significant effects on shrub photosynthesis or growth (Wertin et al., 2015, 2017). Figure 10.5, illustrates carbon cycle interactions and feedbacks associated with multiple climate change factors. Furthermore, changing seasonality of precipitation events, as well as more extreme weather conditions, are expected to affect carbon cycling increasingly more in the future (Knapp et al., 2008b).
Nutrient limitation may reduce the potential for CO2 fertilization in grasslands, especially over decadal timescales (see Figure 10.5). For example, a long-term experiment in a nutrient-poor grassland in Minnesota revealed that elevated CO2 effects on NPP were dependent on soil nitrogen availability and experiment duration. During the first 3 years of the experiment, elevated CO2 stimulated aboveground biomass by 11% and was not contingent on nitrogen availability, but over the longer term (4 to 13 years), the biomass response to elevated CO2 increased by up to 20% with added nitrogen fertilizer (Reich and Hobbie 2013). However, in the coming decades, elevated temperature may enhance nitrogen availability, as shown by Mueller et al. (2016). Moreover, increasing nitrogen deposition will stimulate NPP, up to a threshold, and GHG emissions also may follow a similar nonlinear response to nutrient loading (Gomez-Casanovas et al., 2016). Interacting effects of multiple global change factors still represent a large source of uncertainty in predicting carbon cycle responses (Norby and Luo 2004).
10.3.4. Trends and Climate Feedbacks from Soil Carbon Cycling
The effect of climate change on the stability of carbon in SOM pools is one of the largest sources of uncertainty in projections of climate-carbon interactions (Heimann and Reichstein 2008) because these pools are large and vulnerable to climate change (Davidson and Janssens 2006; see Key Finding 3). In grasslands, decomposition of roots is thought to drive SOM accumulation (Jackson et al., 1996; Jobbagy and Jackson 2000), so processes affecting belowground productivity are likely to affect soil carbon storage (see Figure 10.5). The importance of impacts from aboveground inputs compared to those from direct inputs via root production depends on climate, soil type, and plant species (Sanderman and Amundson 2008). Therefore, grassland species composition and productivity, both above and below ground, and their responses to climatic and land-use changes are key determinants of soil carbon storage. SOM decomposition rates vary with temperature and moisture and can be affected by plant-microbe interactions (van Groenigen et al., 2014) via nutrient uptake processes (Nie and Pendall 2016).
Soil Carbon Responses to Altered Precipitation. Precipitation is the most important climate driver of productivity in grasslands (Knapp and Smith 2001) and is likely to influence carbon storage in soils over longer timescales, via mechanisms related to both plant inputs and decomposition losses (see Figure 10.5). A meta-analysis indicated that soil carbon content increased in response to both reductions and additions of moisture in grasslands (Zhou et al., 2016). Experimentally increased precipitation likely enhanced soil carbon pools via the stimulation of biomass inputs, whereas reduced precipitation may have enhanced the soil carbon pools by reducing SOM decomposition rates as well as by increasing allocation to root biomass production (Zhou et al., 2016).
Soil Carbon Responses to Warming. Earth System Models (ESMs) assume that warming will stimulate SOM decomposition at an exponential rate, leading to potentially strong positive feedbacks to climate change (Figure 10.5; Davidson and Janssens 2006). Experimental evidence of this assumption has been accumulating from numerous individual studies worldwide (Luo 2007). A recent synthesis of warming-experiment results confirms that SOM is vulnerable to warming and indicates that the magnitude of carbon loss depends on initial carbon stocks (Crowther et al., 2016). This study also showed that deserts and arid grasslands, with lower soil carbon pools, are less vulnerable to warming than colder ecosystems. A reduction in decomposition rates with warming-induced soil desiccation could potentially explain these results (Pendall et al., 2013).
Using results from field experiments to inform model parameters is a powerful way to reduce uncertainties, constrain the models, and enhance modeling tools to extrapolate results more broadly. Data from a 9-year warming experiment in tallgrass prairie were assimilated into a biogeochemistry model to demonstrate that soil carbon pools would decrease over the coming century (Shi et al., 2015). This study confirms that carbon in productive grasslands like the tallgrass prairie in Oklahoma can be vulnerable to warming, in part because of the resulting increased decomposition of a large, partially protected soil carbon pool. Key uncertainties were related to the mismatch between the long-term residence time of the large, recalcitrant soil carbon pool and the duration of the experiment (Shi et al., 2015).
Soil Carbon Responses to Rising CO2 and Interactions with Multiple Drivers. While rising atmospheric CO2 concentrations can stimulate grassland productivity above and below ground, especially in combination with warming (Mueller et al., 2016), increased productivity has not necessarily translated into increased soil carbon storage (Luo et al., 2006). A meta-analysis revealed that carbon inputs to grasslands increased by 20% with experimentally increased CO2, but this increase was accompanied by a 16.5% increase in the decomposition rate constant (van Groenigen et al., 2014). The “priming effect” that stimulates SOM decomposition may be caused by the increased microbial activity caused by increased belowground carbon inputs (Carney et al., 2007) and soil moisture (Pendall et al., 2003), and this effect may be “widespread and persistent” (van Groenigen et al., 2014). A simulation model calibrated to realistic field conditions in semiarid Wyoming grassland predicted that soil carbon would decrease with elevated CO2 and increase with warming, because of indirect effects mediated by soil moisture (Parton et al., 2007). However, the importance of interactive effects of multiple climate changes in predictions of long-term soil carbon storage still needs to be confirmed with field results.
Few field experiments have been conducted that combine two or more climate drivers over a long enough duration to evaluate soil carbon responses (Luo et al., 2011), making realistic predictions of soil carbon sequestration challenging. A recent meta-analysis failed to uncover significant changes in soil carbon with the combined effects of elevated CO2 and temperature, although belowground (i.e., root) production was significantly stimulated (Dieleman et al., 2012). While synthesis studies and meta-analyses are useful for discovering general patterns, they cannot distinguish mechanisms underlying these patterns. Major uncertainties in soil carbon storage and ecosystem carbon cycling remain because there are too few long-term, multifactor climate manipulation experiments to constrain mechanisms, feedbacks, and interactive effects among global change drivers.
See Full Chapter & References